This article is made available with the permission of The North Carolina Academy of Science for research purposes only. It is not to be used for commercial activities or reproduced without written permission from the NCAS.
The Journal of the Elisha Mitchell Scientific Society, 103(1),1987, pp.1-13
THE AGE AND TROPHIC HISTORY OF LAKE WACCAMAW, NORTH CAROLINA
J. C. STAGER and L. B. CAHOON
Department of Biological Sciences, University of North Carolina at Wilmington, Wilmington, NC 28403
Abstract: Sediment cores were collected in Lake Waccamaw, a Bay Lake in southeastern North Carolina. The cores were analyzed for information that might be relevant to concerns about the age and trophic status of the lake which is home to several endemic species. Analysis of the diatoms and sponge spicules in the sediments revealed that the lake has always been nearly eutrophic. Analyses of sediment carbon and phosphorus content also support this view. The sediment record indicates that Lake Waccamaw has had periods of shallowing followed by deeper phases. Our interpretation of the geologic history of the Lake Waccamaw area, the sediment record, and the relevant data of others is that Lake Waccamaw is a relatively young lake, probably around 15,000 years old or less. The presence of endemic fauna in the lake is probably a result of the lake's unique chemistry and productivity and not the result of a long period of reproductive isolation from other populations.
Key Words: sediment; eutrophication; endemism; diatoms.
INTRODUCTION
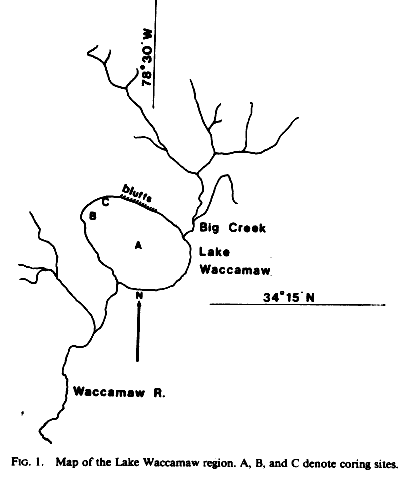
Lake Waccamaw (34o17'N, 78o30'W; Fig. 1) is the largest of the Bay Lakes on the North Carolina coastal plain, with a typically elongated oval shape oriented with its long axis northwest to southeast. The term "Bay" Lake stems from the abundance of bay trees (Magnolia virginiana L., Gordonia lasianthus Ellis, and Persea spp., Frey, 1949) growing in the numerous swampy oval depressions on the Carolina coastal plain. These may represent former lake basins. Many explanations have been advanced for the origin and orientation of the Carolina Bay Lakes and swampy depressions (the 'Carolina Bays"), including meteor impact and limestone dissolution (Melton and Schriever, 1933; Johnson, 1942; Prouty, 1952; Livingstone, 1954; Schriever, 1956; Thom, 1970), but at present wind and wave action are most widely considered to be responsible for the initiation and shaping of the Bays (Kaczorowski, 1977).
Lake Waccamaw is unique among the Bay Lakes for its exceptionally high pH and alkalinity (pH 6.8-7.1, alkalinity 12.0 mg/liter; Weiss and Kuenzler, 1976), in addition to its unusually large size (3,618 ha; Casterlin et al., 1984). Its unique chemistry is at least partially due to the dissolution of calcareous formations that Underlie most of the North Carolina coastal plain, but not the other Bay Lakes. These calcareous deposits form steep bluffs along the north shore of the lake. Finally, Lake Waccamaw is noted for being the only Bay Lake known to support endemic fishes and invertebrates (Hubbs and Raney, 1946; Frey, 1951; Fuller, 977; Casterlin et al., 1984; Porter, 1985).
Recent heavy development of the lake shores has led to concern over potential eutrophication problems that might threaten the water quality of the lake and the existence of its endemic fauna (Casterlin et al., 1984). There is also local concern that the lake may be silting in rapidly as a result of recent human activity along the lake shores and elsewhere in the drainage basin. The work summarized in this paper was undertaken in response to these concerns and was designed to address the following questions: 1) How old is the lake, and how may its environmental history have affected the evolution of its endemic fauna? and 2) What inferences can be drawn about the trophic history of Lake Waccamaw from its sediments, especially with reference to its present trophic status?
SETTING
Lake Waccamaw has a broad, flat bottom of gyttja (mud) and peat encircled by sandy shallows and submerged sandy terraces that extend up to 500 m offshore (Frey,1949). A natural sand bar, which extends along the northwest shore between a fringing cypress (Taxodium ascenclens L.) swamp and the main lake, was supplemented with spoil dredged to form a neighboring canal in 1946 and heavily developed with private residences (John McNeill, pers. commun.). Big Creek delivers tea-colored water from the large cypress and gum swamp at the northeast corner of the lake, and the Waccamaw River emerges from the southern shore. A dam at the outlet built in 1943 now regulates lake levels which used to fall by as much as a meter during dry spells occasionally exposing the sand terraces.
Emergent macrophytes are abundant in the sandy shallows; maidencane (Panicum spp.) is most common near the outlet, and beds of Nuphar and Panicum are widespread along the north shore. Offshore there is a heavy growth of Nitella sp. and Najas on or near the bottom of the lake, much of it covered with green crusts of Spongilla lacustris L.
Benthic microalgae are most abundant at the base of the calcareous bluffs on the north shore of the lake. In May 1986, the littoral sands near the bluffs were bright green with cyanophytes and chlorophytes. The summer phytoplankton community is thought to consist primarily of Oscillatoria, Scenedesmus, and Chlorella (Casterlin et al., 1984), but Melosira valves are also abundant in the offshore gyttja and Botryococcus sp. (B. braunii Kutzing) was common in the plankton in 1985. Strong northwest and southwest winds often churn sediments into the water column probably suspending benthic algae into the plankton whenever the wind exceeds 8 m/sec.
Frey (1950, 1953, 1955a) cored Lake Waccamaw and some of the other Bay Lakes to study their pollen records. The core he collected from Lake Waccamaw consisted of approximately 1 foot of gyttja on top of about 2 feet of "dark sandy silt." Below these soft sediments Frey's coring apparatus was stopped by a dense, sandy blue clay that was practically devoid of pollen. He interpreted the nine pollen subsamples taken from the core to represent a sequence of forest succession passing from pine-spruce through broadleaf to pine forests. On the basis of his interpretation of the pollen record it is reasonable to conclude that the base of the sandy silt layer is late Pleistocene or early Holocene in age (15,000-10,000 yr B.P.) and that cool, moist Pleistocene climates on the coastal plain gave way to warmer, drier conditions during the early Holocene.
Whitehead (1981) studied the pollen record of Rockyhock Bay, N.C., and developed a chronology of environmental events for the northeastern coastal plain of North Carolina. In that region boreal forests gave way to deciduous forests around 10,000 years ago, and a drop in water tables between 7,200 and 5,000 years ago led to the deposition of peat in Rockyhock Bay. The most recent Holocene stage seems to have been established on the coastal plain of North Carolina about 4,000 years ago.
MATERIALS AND METHODS
Three sediment cores were collected from Lake Waccamaw in the spring of 1985. The first core was collected in ca. 3 m of water at the center of the lake (core A: Fig. 1), another was taken in 2 m of water at the northwest end of the lake about 100 m from the public landing (core B), and the third was taken in 0.5 m of water next to the sand bar at the northwest end of the lake (core C). At the last site (core C) the core pipe was initially stopped in peat at a depth of 1m, but further penetration was effected by forcing water down the pipe (piston removed) with a garden hose; slurry emerging from the top of the pipe was collected and the depth of the bottom of the pipe was recorded for each slurry sample.
Geochemical analyses that we used to make inferences about the trophic history of Lake Waccamaw were conducted on subsamples of core A, taken at l-cm intervals. Core A was selected for detailed study because, being from the center of the lake, it was least likely to be influenced by shoreline processes and most likely, therefore, to give a representative picture of the lake's history. Analyses of core A included visual observations of sediment characteristics, water content, percent weight loss on ignition (LOI), organic carbon content, and total phosphorus content. Percent water content was determined after drying the sediment at 60oC for 48 hours, and LOI was determined after ignition at 550oC for 24 hours. Percent organic carbon was determined on dried subsamples using the sulfuric acid-dichromate wet ashing method of Gaudette et al. (1974), and total phosphorus content was determined on dried subsamples using the perchloric acid-nitric acid method of Strickland and Parsons (1972).
Core A was also subsampled at l-cm intervals for diatom analysis. Subsamples were digested in hydrogen peroxide and mounted on glass slides for scanning at 1,000 x . At least 150 diatom valves were counted for each subsample. Diatom identifications and ecological interpretations were based primarily upon Hustedt (1930, 1949), Patrick and Reimer (1966, 1975), Cholnoky (1968), Lowe (1974), Davis and Norton (1978), Gasse (1975, 1980), and Simola (1983).
Surface sediments were collected from 20 sites in and around Lake Waccamaw, including cypress swamps, streams, and other Bay Lakes. Surface samples were scanned at 450 x in order to determine the principal constituents of the modern diatom flora in the area. Measurements of pH were made at most collecting sites with a Cole Parmer pH Wand in May 1986.
RESULTS
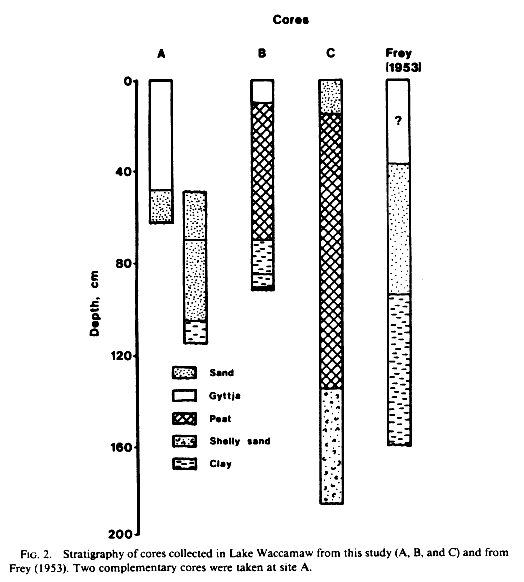
Visual examination of core A revealed three obvious sediment types in the core which was 63.5 cm in length (Fig. 2). The uppermost 20 cm consisted of dark brown gyttja with a small proportion of fine quartz sand. The section between 20 cm and 45 cm also consisted of gyttja but was free of sand. Quartz sand darkened by organic material dominated the core below 49 cm and was interleaved with the overlying gyttja between 45 cm and 49 cm. Fingers of gyttja penetrated the upper few centimeters of the sand layer suggesting the former presence of rooted or burrowing organisms. A second core taken from the same area as core A showed that the sandy layer was underlain by a dense blue clay that was devoid of pollen and siliceous microfossils.
Core B contained four obvious sediment types (Fig.2). Beneath a 7.5-cm veneer of dark brown gyttja lay 62 cm of fibrous brown peat, 16 cm of grey sandy silt, and 5.5 cm of yellow-grey sandy silt. Only the gyttja and peat revealed diatoms or pollen in wet mounts. The upper layer of gyttja could represent the time period since the northwestern cypress swamp was cut off from the main body of the lake by the sand bar as this is when export of plant detritus to this portion of the lake would have been reduced. No date was available for core B, but 7.5 cm of gyttja in Lake Waccamaw could represent 500-1,000 years if Frey's pollen chronology provides an accurate estimate of sedimentation rates.
Core C ended in fibrous brown peat at a depth of ca. 1 m; the peat was blanketed by 15 cm of fine quartz sand (Fig. 2). When the core pipe and hose were pushed below the 135-cm level a sandy shell hash containing sea urchin spines and buff-colored silt emerged from the top of the pipe. The sandy shell hash appeared to extend at least 50 cm below the level of the peat.
Analyses of percent water content and LOI supported our initial decision to subdivide core A into three stratigraphic sections (Fig. 3). Values of both measurements were clearly higher in the middle portion of the core than in the upper section and were lowest in the sandy basal layer.
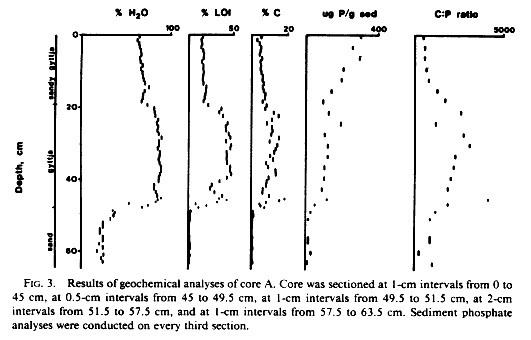
Analyses of percent organic carbon in core A showed a pattern similar to those of water content and LOI (Fig. 3). Organic carbon content of the middle section of the core was roughly twice as high as that of the upper section and much higher than that of the sandy basal layer.
Analyses of total phosphorus content in every third sediment subsample revealed a distribution that was very different from that of organic carbon (Fig. 3). Phosphorus content in the upper section of the core was higher than that of the rest of the core causing carbon: phosphorus (C:P) atomic ratios to be much higher in the carbon-rich middle section of the core than elsewhere.
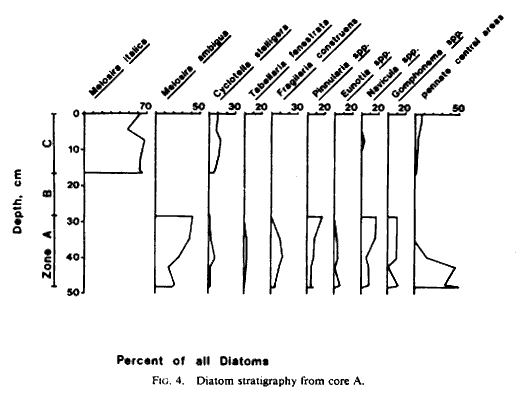
Diatoms were abundant in the upper 48 cm of core A but were absent from the sandy basal layer, probably as a result of mechanical degradation in the coarse sediments. The diatom record of core A was subdivided into three zones (Fig. 4) as follows:
1) Zone A (48.0-28.5 cm) was dominated by fragments of various benthic pennate diatoms belonging to such genera as Navicula and Pinnularia. Melosira ambigua Grun increased relative to other diatoms in the upper part of zone A; it is commonly found in the plankton of eutrophic lakes in the tropics and temperate zones, especially but not exclusively in alkaline waters. Other common diatoms were Cyclotella stelligera Cleve (planktonic and/or benthic, alkaline to circumneutral, eutrophic to oligotrophic waters) and Fragilaria construens Lyngbye (benthic and facultatively planktonic, alkaline eutrophic waters). Sponge spicules, mostly fragmented, increased in abundance relative to diatoms in the upper portion of zone A along with M. ambigua. The spicules closely resembled those of Spongilla lacustris, but positive identification was not considered feasible on the basis of megascleres alone.
2) Zone B (28.5-16.5 cm) contained many broken sponge spicules but very few diatoms. Those valves that did appear in zone B were heavily corroded and fragmented, but it was possible to identify a small number of Pinnularia, Navicula, Eunotia, and Melosira spp. in the lower and upper portions of the zone. Melosira was represented by M. ambigua in the lower subsamples and by M. italica Grun (planktonic or benthic in alkaline, eutrophic waters) in the upper subsamples. Because of the low numbers of valves, the diatom assemblage of zone B was not included in Figure 4.
3) Zone C ( 16.5-0.0 cm) was dominated by M. italica, which occurred in small and large forms similar to illustrations of M. italica var. italica and M. italica var. valida (Hustedt, 1930). Next in abundance was C. stelligera; pennate diatoms were relatively rare, and sponge spicules varied in abundance. Stephanodiscus sp. cf. S. dubius Ehrenberg occurred in very small numbers in zone C. Although the species identification was not certain, the genus itself is best represented in alkaline, relatively eutrophic waters, and its presence in Lake Waccamaw's sediments was not surprising.
The gyttja at the top of core B contained the same diatom assemblage as the upper section of core A: M. italica varieties and C. stelligera were the dominant taxa. Below 7.5 cm the fibrous peat contained smaller numbers of diatoms, most of which were benthic, pennate forms, such as Eunotia spp. and Pinnularia spp. typical of acidic conditions, although a small proportion of M. italica persisted throughout the peat section. Sponge spicules were abundant in the basal sandy silts, but diatoms were rare. Only a few corroded pennate valves were found in the silts, suggesting that the sandy silts of core B were contemporaneous with the zone B gyttjas offshore.
Surface sediments collected from offshore stations in Lake Waccamaw contained large numbers of M. italica valves, along with C. stelligera and large, mostly broken sponge spicules. Although the diatom flora was clearly dominated by Planktonic or semi-planktonic forms, a small proportion of primarily benthic diatoms belonging to the genera, Achnanthes, Pinnularia, and Navicula was present also in the sediments. Closer to shore, the sediments became grittier and eventually gave way to sands. The relative proportions of planktonic and benthic diatoms reversed as water depths decreased and the sediments became coarser. Cyanophytes and chlorophytes were found attached to many of the littoral sand grains.
The weed-choked canal behind the sand bar on the northwest shore of the lake yielded many Eunotia, Pinnularia, Navicula, and Gomphonema; centric planktonic diatoms were absent. Big Creek (pH 6.0) supported a similar flora along with filamentous cyanophytes. Eunotia was the dominant diatom genus in the swampy streams feeding Big Creek (pH ranges 6.1-6.6). The littoral sediments of three Bay Lakes in nearby Bladen County were also sampled for comparative purposes. Jones Lake (pH 3.1-4.8, alkalinity 0.0-0.1 mg/liter; Weiss and Kuenzler, 1976) had filamentous and globular cyanophytes in its sandy sediments along with Eunotia, Pinnularia, Navicula, and Surirella. White Lake (pH 4.6-4.8, alkalinity 0.1-0.5 mg/liter; Weiss and Kuenzler, 1976) was much clearer than Jones Lake, and its littoral sands were devoid of algae except for a few Pinnularia, Navicula, and some filamentous chlorophytes. Singletary Lake (pH 3.2-4.6, alkalinity 0.0-0.1 mg/liter; Weiss and Kuenzler, 1976) was the only Bladen County lake in this survey that supported planktonic diatoms. Asterionella cf. A. formosa Hassall (eutrophic waters) dominated the diatom flora in the sandy littoral sediments which also included Pinnularia, Eunotia, and Navicula spp.
The results of the survey of modern diatom floras suggest that Lake Waccamaw's unique chemistry has contributed to the development of an equally unique alkaliphilic planktonic diatom community in a region dominated by acidic swamps and lakes. The occasional Eunotia or Pinnularia encountered offshore in Lake Waccamaw probably represents diatom transport from the shore and surrounding swamps or resuspension of sediments by wind-driven mixing. Offshore the sediments are fine and highly organic, rich in planktonic diatoms; inshore they become sandy, relatively inorganic, and richer in benthic microalgae than in planktonic ones.
DISCUSSION
The patterns of water content and loss on ignition in core A do not necessarily tell much about the trophic history of the lake. Water content and LOI were lower in the sandy section than in the gyttjas because of the nature of the two sediment types: coarse quartz grains exclude more water and organic matter than fine silty sediments. However, it may be possible to deduce the history of lake depths from these data as the shallows of Lake Waccamaw today are underlain by sands that give way to gyttjas offshore. It appears likely that the sandy portion of core A represents a period of shallowing, followed by a deeper or calmer period of gyttja deposition that continues today. If Frey's chronology is applied to core A, the end of the lake's shallow phase would have occurred around 5,000 years ago, corresponding to Frey's proposed onset of cooler, moister conditions (Frey, 1953, 1955a). Whitehead's (1981) chronology would reduce the age of the gyttja to ca. 4,000 years.
Phosphorus depletion of the Lake Waccamaw sediments suggested by the high carbon: phosphorus atomic ratios in the middle section of core A might have been caused by intense scavenging of phosphorus by macrophytes over time. Nitella is abundant at the coring site today, and rooted vegetation may have been widespread in the past. Benthic diatoms were more numerous in zone A than in zone C, which might indicate that the lake was shallower while the zone A gyttjas were being deposited. The poor preservation of diatoms in zone B is enigmatic at present, but it might reflect intense reworking of the sediments by currents or organisms; the abundance of spicules in zone B may reflect shallow or clear lake conditions. The data at hand argue strongly for biological uptake of phosphorus in Lake Waccamaw sediments since even the lowest carbon: phosphorus ratios in core A are much higher than the Redfield ratio for living matter (roughly 105: 1).
An alternative explanation for phosphorus depletion of the Waccamaw sediments is the occurrence of anoxic conditions, which would create a reducing environment that would in turn cause solubilization of phosphorus as ferrous phosphates. Anoxia in the water column would imply highly eutrophic conditions, but even under such conditions anoxia is unlikely to persist for long in such a shallow lake with a long fetch that facilitates frequent wind mixing. For now, phosphorus scavenging by benthic organisms appears to be the most reasonable explanation for phosphorus depletion in Lake Waccamaw sediments.
The diatom record of core A indicates that Lake Waccamaw has always been incipiently eutrophic; traces of cyanophytes were also visible on the early Holocene sand grains as on the green littoral sands of today. The planktonic diatom floras of zones A and C were both typical of productive lakes in the temperate zone, although zone A probably reflected a shallower lake that might have been more thoroughly mixed than that of today. Natural phosphate loading and high alkalinity from the dissolution of marine formations have made the lake chemically unique among the Bay Lakes and able to support a diatom flora dominated by alkaliphilic diatoms through most, if not all, of the Holocene. The naturally productive nature of Lake Waccamaw suggests that human inputs of phosphorus or nitrogen could force it quickly to hypereutrophy; the littoral sands along the north shore are already green with benthic microalgae and associated filamentous algae, and rooted macrophytes are said by local residents to be on the increase. There is, however, no evidence that the lake is filling in rapidly; it seems to have been accumulating sediments offshore as slowly as 0.05-0.10 mm per year for the last 10,000 years or so. If anything, the diatom record suggests that the lake has actually been getting deeper during the late Holocene.
Recent studies conducted on the regional geology of the Carolina coastal plain (Zullo and Harris, 1979) suggest that the Waccamaw and Cape Fear drainages formed one unit flowing to the southwest before crustal warping disrupted river patterns during the Waccamaw-Canepatch Uplift between 75,000 and 32,000 years ago. The Socastee marine transgression, about 32,000 years ago, may have submerged the site of modern Lake Waccamaw. If this geologic scenario is correct then Lake Waccamaw must be less than 32,000 years old.
The coastal plain around Lake Waccamaw [310k] is underlain by marine sands, marls, and clays of Pleistocene age and older (V. A. Zullo, pers. commun.). The bluffs on the north shore of the lake are Plio-Pleistocene and Cretaceous in age (S. Snyder, pers. commun.). It is reasonable to conclude that the blue clay underlying the sand layer in core A belongs to a marine formation as does the shell hash in core C. Unfortunately, the lack of microfossils in the blue clay makes it difficult to determine its origin. Whether it was deposited in fresh or salty waters, the density, low water content, and sharp contact with overlying sands suggest that the clay was exposed to drying and erosion. If the clay-sand interface represents an erosional unconformity, and Frey's chronology (Frey, 1955a) is accurate, then the existence of Lake Waccamaw can only be substantiated for the last 15,000 years. With an age of 15,000 years, it would be one of the youngest of the Carolina Bay Lakes. Singletary Lake, which lies inland from Waccamaw among the Bladen County Lakes, has yielded a pollen record that extends beyond the range of radiocarbon dating (ca. 50,000 years) (Frey, 1953, l955a,b). Even if the blue clay were deposited in the lake immediately prior to the sands, it seems very unlikely that Lake Waccamaw predates the Waccamaw-Canepatch Uplift, or even the Socastee Transgression of 32,000 yr B.P.
The unusually large size of Lake Waccamaw, its unique position astride a major river, and the presence of marine bluffs along the north shore of the lake argue strongly for a tectonic initiation of the lake near the close of the Pleistocene; wind and wave action subsequently led to the development of its elliptical shape as was probably the case with the other Carolina Bays (Kaczorowski, 1977). The sand rim, which can be observed easily at the northwest corner of the lake, probably represents the remnant of an earlier shoreline that developed under moist Pleistocene conditions when water tables on the coastal plain were higher than today (Whitehead, 1981).
Sand rims are one of the trademarks of the Carolina Bays (Frey, 1949; Prouty, 1952), but far less attention has been devoted to the nature of their origin than to that of the elliptical basins and their orientations. It is likely that the Bays contained open water at some point in their development, probably during the Pleistocene moist phase. Since modern Carolina winters occasionally bring freezing temperatures and since glacial Pleistocene winters were colder than winters today (Whitehead, 1981), it is likely that the Pleistocene Bay Lakes froze partially or completely each year. Collision of wind-driven ice with the sandy shore combined with thermal expansion of ice on completely frozen lakes could have left substantial ice-push ridges around the perimeters of the lakes with the strongest development on the down-wind shores; in fact, sand rims around Carolina Bays are best developed on the southeast, or downwind, sides of the basins (Prouty, 1952). The formation of ice-push ridges has been well documented for lakes at higher latitudes, including those with rocky and gravel-strewn shorelines that would be more resistant to ice action than loose sands (Leverett and Taylor, 19 l 5; Washburn, 1947; Hume and Schalk, 1964; Hult, 1968; Dionne and Laverdiere, 1972; Embleton and King, 1975). Concentric and intersecting sand rims in some Carolina Bays (Prouty, 1952) may therefore reflect transgressions or oscillations in lake levels.
We propose that Lake Waccamaw is a relatively young lake that probably formed through crustal warping near the close of the Pleistocene. lf our hypothesis is correct, then the fact that Lake Waccamaw is the only Carolina Bay Lake known to support endemic fauna may be surprising as it is often assumed that old age is a prerequisite for the development of endemic species. Evidence to the contrary has come from studies on Lake Victoria, East Africa, where a large species school of cichlid fishes has developed in a lake that may be less than 15,000 years old (Stager et al.,1986; B. Rosendahl, pers. commun.,1986). Lake Nabugabo, isolated from Lake Victoria by a drop in lake levels during the late Holocene, also contains endemic cichlids that would necessarily have evolved in less than 4,000 years (Greenwood, 1965). The other features that make Lake Waccamaw unique among the Bay Lakes, such as its high alkalinity, calcium content, productivity, and its large size, may have more to do with the onset of speciation than age alone.
The stratigraphy of core B suggests that the rate of peat deposition at the northwest end of the lake was greatly reduced between 500 and 1,000 years ago. Whether the peat in the core represents a cypress swamp or a sludge fan from a nearby swamp, its succession by gyttja appears to have been caused by a recent nse in lake levels which pushed back the edge of the northwest swamp and led to the growth of a sand bar barrier. This hypothesis is supported by earlier work done on cypress trees isolated from shore in Lakes Waccamaw and Singletary and White Lake which suggested that the lakes have been expanding for "at least the past 500 years" (Frey, 1954). The presence of sand in the upper 20 cm of core A may reflect an increase in the transport of sands from the shore during lake expansion or, on the other hand, an offshore retreat of sandy shallows during a shrinking phase. The possibility that the true surface of the soft gyttja was disturbed or missed by overdriving the coring apparatus makes it unwise to put too much trust in the uppermost stratigraphy of cores A and B. Further sampling of the mud-water interface will be required to resolve speculations on lake level fluctuations in recent times.
Acknowledgments: The authors would like to thank John McNeill for consultations and the use of his facilities on Lake Waccamaw. R. B. Davis and D. Anderson provided access to their expertise and equipment at the University of Maine at Orono, and the Department of Biological Sciences at UNC-Wilmington provided field support and supplies. This work was also supported by NSF grant RII 8311 486. K. Haberyan identified the Botryococcus, and Ani Stenkvist helped nurture the idea of ice-push ridges. S. Snyder of the Geology Department at East Carolina University identified the rocks from the north shore of Lake Waccamaw. We would like to thank D. G. Lindquist, D. G. Frey, and V. A. Zullo for critical reviews of the manuscript.
REFERENCES CITED
CASTERLIN, M. E., W. W. REYNOLDS, D. G. LINDOUIST, AND C. G. YARBROUGH. 1984. Algal and physioehemical indicators of eutrophication in a lake harboring endemic speeies: Lake Waccamaw, North Carolina. J. Elisha Mitchell Sci. Soe. 100:83-103.
CHOLNOKY, B. 1. 1968. Die Okologie der Diatomeen in Binnengewassern. Cramer-Lehre. 699 pp.
DAVIS, R. B., AND S. A. NORTON. 1978. Paleolimnologic studies of human impact on lakes in the United States, with emphasis on recent research in New England. Pol. Arch. Hydrobiol. 25: 99-115.
DIONNE, J. C., AND C. LAVERDIERE. 1972. Ice-formed beach features from Lake St. Jean, Quebec. Can. J. Earth Sci. 9:979-990.
EMBLETON, C., AND C. A. M. KING. 1975. Glacial and Periglacial Geomorphology. Vol. 2. Edward Arnold, London. 203 pp.
FREY, D. G. 1949. Morphometry and hydrography of some natural lakes of the North Carolina coastal plain: the Bay Lake as a morphometric type. J. Elisha Mitchell Sci. Soe. 65: 1-37.
FREY, D. G. 1950. Carolina Bays in relation to the North Carolina coastal plain. J. Elisha Mitehell Sci. Soe. 66:44-52.
FREY, D. G. 1951. The tishes of North Carolina's Bay Lakes and their intraspecific variation. J. Elisha Mitchell Sci. Soe. 67: 1-44.
FREY, D. G. 1953. Regional aspects of the late-glacial and post-glacial pollen succession of south-eastern North Carolina. Ecol. Monogr. 23:289-313.
FREY, D. G. 1954. Evidence for recent enlargement of the "Bay Lakes" of North Carolina. Ecology 35: 78-88.
FREY, D. G. 1955a. A time revision of the Pleistocene pollen chronology of southeastern North Carolina. Ecology 36:762-763.
FREY, D. G. 1955b. Stages in the ontogeny of the Carolina Bays. Proc. Int. Assoc. Appl. Limnol.12:660- 668.
FULLER, S. L. H. 1977. Freshwater and terrestrial molluscs. Pp. 143-194 in J. E. Cooper, S. S. Robinson, and J. B. Funderberg (Eds.), Endangered and Threatened Plants and Animals of North Carolina. N.C. State Museum of Natural History, Raleigh.
GASSE, F. 1975. L'evolution des lacs de l'Afar Central du Plio-Pleistocene a Actuel. Ph.D. thesis, Univ. of Paris. 403 pp.
GASSE, F. 1980. Les diatomees lacustres Plio-Pleistocenes du Gadeb (Ethiopie): systematique, palaeoecologie, biostratigraphie. Rev. Algol. 3. 249 pp.
GAUDETTE, H. E., W. R. FLIGHT, L. TONER, AND D. W. FOLGER. 1974. An inexpensive titration method for the determination of organic carbon in recent sediments. J. Sedim. Petrol. 44:250- 253.
GREENWOOD, P. H. 1965. The cichlid fishes of Lake Nabugabo, Uganda. Bull. Brit. Mus. Nat. Hist. Zool. 12:315-357.
HUBBS, C. L., AND E. C. RANEY. 1946. Endemic fish fauna of Lake Waccamaw. Misc. Publ. Mus. Zool. Univ. Mich., 65. 30 pp.
HULT, J. 1968. Some Aspects of the Shore Formation of Lake Lylykkaanjarvii, Finland. Fennia 97. 22 pp.
HUME, J. D., AND M. SCHALK. 1964. The effects of ice-push on Arctic beaches. Am. Jour. Sci. 262: 267-273.
HUSTEDT, F. 1930. Bacillariophyta (Diatomae). In Pascher, Die Susswasser-Flora Mitteleuropas. Zweite Auflage 10. G. Fischer, Jena. 466 pp. 1949. Susswasser Diatomeen aus dem Albert-National Park in Belgisch-Kongo. Exploration du Parc National Albert. Mission H. Damas (1935-1936). Brussels, Inst. des Parcs Nationaux du Congo Belge. Fasc. 8. 199 pp.
JOHNSON, D. W. 1942. The Origin of the Carolina Bays. Columbia Univ. Press, New York. 341 pp.
KACZOROWSKI, R. T. 1977. The Carolina Bays: a comparison with modern oriented lakes. Tech. Rpt. 13-CRD. Coast. Res. Div., Univ. S.C.
LEVERETT, F., AND F. B. TAYLOR. 1915. The Pleistocene of Indiana and Michigan, and the History of the Great Lakes. Geol. Soc. Am. Monogr. 53.
LIVINGSTONE, D. A. 1954. On the orientation of lake basins. Am. Jour. Sci. 252:547-554.
LOWE, R. L. 1974. Environmental Requirements and Pollution Tolerance of Freshwater Diatoms. Envir. Prot. Agency, EPA-670/4-74-005. 334 pp.
MELTON, F. A., AND W. SCHRIEVER. 1933. The Carolina Bays --are they meteor scars? Jour. Geol. 41 :52-66.
PATRICK, R., AND C. W. REIMER. 1966. The Diatoms of the United States, Vol. 1. Monogr. Acad. Nat. Sci. Philadelphia, 13. Livingston, U.S.A. 688 pp.
PATRICK, R., AND C. W. REIMER. 1975. The Diatoms of the United States, Vol. 2. Monogr. Acad. Nat. Sci. Philadelphia, 13. Scutter House Lititz, U.S.A. 213 pp.
PORTER, H. J. 1985. Rare and Endangered Fauna of Lake Waccamaw, North Carolina Watershed System: Molluscan Census and Ecological Interrelationships. North Carolina Wildlife Resources Commission. 187 pp.
PROUTY, W. F. 1952. Carolina Bays and their origin. Geol. Soc. Am. Bull. 63:167-224.
SCHRIEVER, W. 1956. Discussion of "Were the Carolina Bays oriented by gyroscopic action?" Am. Geophys. Union Trans. 37: 112-115.
SIMOLA, H. 1983. Limnological effects of peatland drainage and fertilization as reflected in the sediments of a deep lake. Hydrobiologia 106:43-57.
STAGER, J. C., P. N. REINTHAL, AND D. A. LIVINGSTONE. 1986. A 25,000 year history for Lake Victoria, East Africa, and some comments on its significance for the evolution of cichlid fishes. Freshw. Biol. 16:l5-l9.
STRICKLAND, J. D. H., AND T. R. PARSONS. 1972. A Practical Handbook of Seawater Analysis. Fish. Res. Bd. Can., Ottawa. 310 pp.
THOM, B. G. 1970. Carolina Bays in Horry and Marion Counties, South Carolina. Geol. Soc. Am. Bull. 81:783-814.
WASHBURN, A. L. 1947. Reconnaisance Geology of Portions of Victoria Island and Adjacent Regions, Arctic Canada. Am. Geol. Soc. Mem. 22.
WEISS, C. M., AND E. J. KUENZLER. 1976. The Trophic Status of North Carolina Lakes. Rpt. 119, Water Resources Res. Inst., Univ. N.C.
WHITEHEAD, D. R. 1981. Late-Pleistocene vegetational changes in northeastern North Carolina. Ecol. Monogr. 51 :451-471.
ZULLO, V. A., AND W. B. HARRIS. 1979. Plio-Pleistocene crustal warping in the outer coastal plain of North Carolina. Pp. 31-40 in G. R. Baum, W. B. Harris, and V. A. Zullo (eds.), Structural and Stratigraphic Framework for the Coastal Plain of North Carolina. Carolina Geol. Soc. and Atlantic Coast. Plain Geol. Assoc.
Received 22 January 1987
![coastal plain around Lake Waccamaw [310k]](WACTER.GIF){kind=link}